
zdarzało się niejednokrotnie, że stan jakiejś cechy lub sekwencji nukleotydów w
DNA zmieniał się, a potem wracał do formy wcześniejszej. Zjawisko to nazywamy
rewersją – wtórnym wykształceniem jakiegoś parametru, który został w trakcie
ewolucji utracony lub ewentualnie znacznie zmarginalizowany. Badaczom historii
naturalnej udało się zidentyfikować wiele różnych, często ciekawych i
zaskakujących rewersji (także u człowieka). Oto niektóre z nich.

ciekawych przykładów, chciałbym wyjaśnić kilka kwestii. O stanie wtórnym i
pierwotnym oraz o rewersji możemy mówić w odniesieniu do różnych momentów w
rozwoju danej grupy. Jeżeli bierzemy pod uwagę kilka kolejnych kladów (czyli
grup rozwojowych), to względem starszych grup rozpatrujemy rewersje u grup
młodszych. Stan jakiejś cechy może być rewersją względem starszego wspólnego
przodka, niekoniecznie względem młodszego i na odwrót. Jest to więc pojęcie
płynne, zależnie od tego, do jakiej cechy, jakiego stanu cechy i jakiej grupy
organizmów będziemy się odnosić.

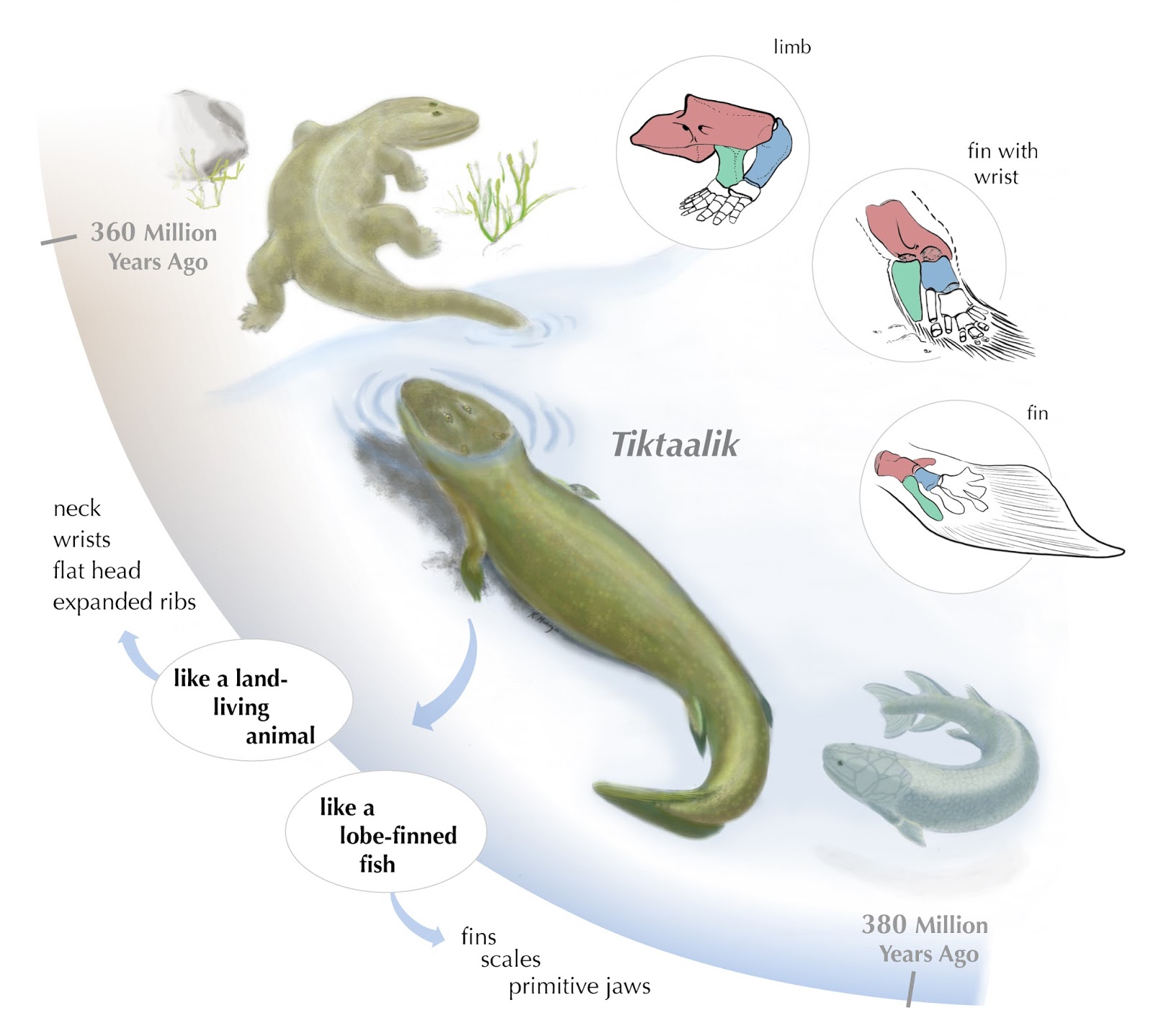
zwierzę, kręgowca, które ma nogi i ręce, a które ewoluowało z beznogiego
przodka, nie będącego jednak rybą, a gadem (czyli wtórnie beznogiego, bo gady
wyewoluowały z płazów, a te wcześniej z ryb). Względem tego przodka obecność rąk
i nóg nie będzie więc rewersją, nie będzie nią też względem najbardziej
pierwotnego w uwzględnianej konfiguracji przodka rybiego (gdyż płetwy, choć są
kończynami, ich budowa i funkcja odbiega od kończyn kręgowców lądowych), ale
będzie rewersją porównując do grupy, która nastąpiła po rybach, a przed
beznogim gadem. W związku z tym to, czy coś jest rewersją zależeć będzie do
tego, do jakiej wcześniejszej grupy przodków się odnosimy.
 |
| (Dawson, Wikipedia) |
przykład możemy przynajmniej częściowo przełożyć na rzeczywisty przypadek w
historii życia na naszej planecie. Węże w wyniku ewolucji pod ziemią utraciły kończyny
przednie i tylne bądź też zachowały je w szczątkowej formie (zrosły im się
także powieki, a ponadto niektóre z nich wykształciły termoreceptory
pozwalające odbierać podczerwień), pochodzą jednak od czworonożnych jaszczurek,
które z kolei wywodzą się od beznogich kręgowców morskich. Podobną sytuacją
mogą poszczycić się marszczelce, czyli beznogie płazy, podobne wyglądem do
robaków, które także w wyniku adaptacji do podziemnego trybu życia utraciły kończyny,
chociaż istnieje debata wśród naukowców nad tym, czy marszczelcowate nie
ewoluowały niezależnie od reszty płazów.

będzie cechował się rewersją, gdyż płetwy u tych zwierząt są de facto kończynami, ale też nie rozumianymi jako te u czworonogów (problem
nazewnictwa ten, co wyżej, czworonogi to po prostu zwierzęta, które
wyewoluowały pierwotnie z wczesnych płazów, samo słowo zaś pochodzi od
Tetrapoda, w języku polskim używanym jako tetrapody), będzie za to rewersją
względem wcześniejszych, morskich kręgowców, jak np. jednego z najstarszych nam
znanych Haikouichthys. Gdyby z kolei
z węży wyewoluowały w przyszłości zwierzęta z wykształconymi kończynami (co jak
najbardziej może się zdarzyć), mielibyśmy realny przykład odzwierciedlający ten
powyższy, hipotetyczny. Warto przyjrzeć się zdjęciom ukazującym szczątkowe nogi
węży.
 |
(Haikouichys, Nobu Tamura, Wikipedia) |
 |
|
Sedmera, D., I. Misek, and M. Klima.
„On the development of Cetacean extremities:
I. Hind limb rudimentation in the
Spotted dolphin (Stenella attenuata).”
|
Pozostańmy jeszcze chwilę przy temacie
kończyn, nóg, rąk i płetw, bo kolejnym przykładem będą walenie. Ich ewolucja to jeden z moich ulubionych tematów – jak
pewnie zauważyliście, czytając poszczególne wpisy. Prawdopodobnie domyślacie
się, co będzie dalej, jeżeli przeczytaliście akapity wyżej, ale bądźmy
oficjalni. Płetwy, które wykształciły się u wielorybów i delfinów (oraz
morświnów) są kończynami przednimi, rękami. Tylne zostały silnie zredukowane
(dając w efekcie nogi szczątkowe) bądź – u niemal wszystkich przypadków –
zanikły. Nie oznacza to, że geny je kodujące zostały utracone, po prostu ich
ekspresja jest epigenetycznie zahamowana, a kończyny tylne u waleni, w
większości przypadków spotkamy jedynie w okresie rozwoju płodowego.
Oczywiście rząd tych ssaków nie
wziął się znikąd lecz, podobnie jak cała reszta kręgowców, od pewnego momentu
miał swój początek u ryb, wobec których wykształcenie płetw – po ich utraceniu
przez stadium płazów, gadów i ssaków lądowych – jest zjawiskiem rewersji. Za
siostrzane przykłady posłużyć nam mogą syreny (w randze rzędu) – reprezentowane
współcześnie jedynie przez diugonie i manaty (a do XVII wieku także przez
ogromne krowy morskie, wytępione przez marynarzy) oraz wymarłe już gady morskie,
przystosowane do środowiska wyłącznie wodnego, które reprezentowane były np.
przez ichtiozaury, bardzo podobne morfologicznie do znanych nam dzisiaj
delfinów.

 |
| Pixabay |
Bardzo ciekawym przykładem jest
także uzębienie waleni, z podrzędu zębowców (zauważcie, że grupa ta dzieli się
na ogromne fiszbinowce, filtrujące wodę w poszukiwaniu pokarmu, za pomocą
fiszbin oraz drapieżne zębowce, do których zaliczają się delfiny, orki,
morświny, narwale, kaszaloty). Zanim przejdziemy dalej, przyjrzyjcie się
najpierw uzębieniu bardziej pierwotnych kręgowców – prymitywnych ryb,
prymitywnych płazów, gadów, prymitywnych, wymarłych ptaków oraz ssaków.
Porównajcie zęby przykładowo psa, z zębami innych zwierząt należących do
wymienionych grup. Co widzicie?
 |
| Gawial, Wikipedia |
ryb, płazów, gadów i ptaków jest jednorodne, homogenne, nie mamy szczególnego
zróżnicowania na różne typy, w przeciwieństwie do zębów ssaków, które dzielą
się ogólnie na siekacze, kły, przedtrzonowce i trzonowce, wyróżniamy je też na
podstawie guzków. Teraz spójrzcie na zdjęcie niżej, przedstawiające uzębienie
waleni zębowych. Jest ono jednolite – zęby są mniej więcej takie same, tego
samego rodzaju, bez różnorodności w występowaniu guzków. Podobnie jak w
przypadku gadów, prymitywnych ptaków i płazów oraz ryb. Ostatni znacznie różniący
się morfologią przodkowie waleni, prakopytne, posiadały uzębienie
charakterystyczne dla reszty ssaków, odróżnicowanie go zaczęło następować wraz
z przystosowywaniem się do środowiska wodnego.

należą do grupy owadów uskrzydlonych, to wykształconych przydatków umożliwiających
aktywne latanie nie posiadają. Wywodzą się jednak z owadów, które posiadały (z
kolei te uskrzydlone ewoluowały z wcześniej bezskrzydłych i stąd ta wtórność),
a ponowne wystąpienie tej cechy po przejściu cechy przeciwnej także jest
klasycznym przykładem rewersji. Być może latające współcześnie chrząszcze (jak
np. biedronka) za kolejne tysiące lat ewolucji staną się owadami wtórnie
bezskrzydłymi. Już teraz jedna para skrzydeł jest u tej grupy zwierząt
przekształcona w pokrywy chitynowe, a niektóre gatunki w ogóle nie są zdolne do
lotu. Przyjrzyjcie się startującej w powietrze biedronce.
możemy także na poziomie molekularnym w postaci mutacji podstawiającej
nukleotyd, który wcześniej został podmieniony na inny. Czyli przykładowo, w
danym miejscu sekwencji DNA mamy ciąg zasad AGCT (adenina, guanina, cytozyna,
tymina), następnie zachodzi mutacja, podstawiająca C na G i nasza informacja
wygląda następująco: AGGT. Po jakimś czasie obserwujemy kolejną zamianę
nukleotydów, z powrotem z G na C w tym samym miejscu – zachodzi rewersja.
Zjawisko rewersji molekularnej wcale nie musi i często nie będzie szło w parze
z rewersją fenotypową (czyli taką, o której pisałem w pierwszych akapitach) w
skali ewolucyjnej, ponieważ wystąpienie danych cech podobnych do wcześniej
utraconych niekoniecznie musi wynikać z odtworzenia pierwotnej sekwencji DNA.

innych pozycjach, niż pierwotnie miało to miejsce, ale przywracające kodowanie pierwotnie
odczytywanego aminokwasu (lub cechy fenotypowej), co możliwe jest dzięki temu, iż kod genetyczny jest
zdegenerowany – kilka kodonów informuje o jednym aminokwasie. Pojedyncze
podmiany nukleotydów często doznają rewersji, natomiast duplikacje lub delecje
(czyli utrata) dłuższych fragmentów sekwencji – bardzo rzadko. Jako ciekawostkę
warto przytoczyć wykorzystanie zjawiska rewersji nukleotydowej w skali
nieewolucyjnej do badania mutagenności w teście Amesa, gdzie bakterie
posiadające mutację uniemożliwiającą syntezę niezbędnego dla nich do życia
aminokwasu traktowane są różnymi mutagenami. Im więcej kolonii bakterii
wyrośnie na pożywkach pozbawionych aminokwasu, którego zmutowane bakterie nie
syntetyzują, tym bardziej mutagenny jest czynnik, którym zostały potraktowane, gdyż
w większej liczbie przypadków zaszła mutacja odwrotna, pozwalająca na powrót
syntetyzować dany aminokwas, tym samym pozwalając bytować bakterii w pożywce go
pozbawionej.

rewersja nie oznacza cofnięcia się ewolucji danego gatunku. Konkretna cecha,
która została przywrócona jest efektem działania doboru naturalnego, jest zatem
dalszą ewolucją przystosowującą populację do środowiska, w którym żyje i w
którym ewoluuje, a to środowisko może w jednym okresie czasu wymuszać, na
przykład, wykształcenie skrzydeł, w innym potrzebę ich posiadania
marginalizować, by następnie znów sprawić, że te staną się przydatne. Mówienie
więc o cofaniu się ewolucji jest błędem merytorycznym, możemy za to stwierdzić,
że jakaś cecha się uwsteczniła czy pojawiła wtórnie – jak to robiłem
wielokrotnie w tym artykule.
Chcesz wesprzeć rozwój mojego bloga? Możesz to zrobić zostając jego patronem tutaj.










Bardzo mnie to zagadnienie interesowało już w szkole, chociaż niewiele się o tym mówiło. Bardzo mi się ten artykuł podobał, fajnie rozwinąłeś ten temat. Pozdrawiam!
dokładnie! Za mało tego co ciekawe uczą w szkołach… ech… niestety taki system 🙂
no nieźle, bardzo rzadko czytam wpisy w tym stylu 😀
lubię wszelkie gady, ale nigdy się nie interesowałam, jak dokładnie ewoluowały.
fajne te nogi szczątkowe
nigdy mnie to jakoś nie interesowało.. ogólnie tematyka w ogóle nie moja, ale ciekawie to opisujesz:)
Ewolucja?Jaka ewolucja?!Przecież bug stworzył świat 4000 lat temu ( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)( ͡° ͜ʖ ͡°)
Bardzo ciekawe jest to co piszesz, to niezwykłe jak wszystko na przestrzeni czasu ulega modyfikacjom jednak ma to jakiś sens i jest według pewnego schematu. 🙂
mnie ciekawią takie zagadnienia ewolucji zmian modyfikacji:) za młodu namiętnie pochłaniałam książki o tym:)
W bardzo ciekawy sposób przybliżyłeś mi temat rewersji 🙂
nie przypuszczałam że wąż kiedyś tak wyglądał
Przyznam się, że od czasu do czasu lubię patrzeć na ludzi i zwierzęta (najczęściej na moją Rubi) i szukać w nich śladu przodków. Czasem to układ ciała w jakiejś konkretnej pozie, czasem cień posiadanych kiedyś kończyn, którymi chciałoby się zamachnąć, ale ich już dawno nie ma. Mam nadzieję, że nie jestem zbyt creepy 😀
Najbardziej spodobała mi się puenta – ewolucja się nie cofa. To jest coś, co gryzło mnie w oczy czy uszy wiele razy. Ewolucja to proces "do przodu". Cofanie się to proces "do tyłu". To tak jakby mówić: "Cofam się do przodu". Oczywiście można określić prędkość do przodu i do tyłu i w ten sposób wyznaczyć wypadkową, ale w biologii raczej nie o to chodzi… :))
Łukaszu, Twoje, niewątpliwie ciekawe, wywody udowodniły mi, jak mało wiem o ewolucji.